
Was genau bringt eine Pflanze zum Blühen? Wir können diese Frage noch nicht abschließend beantworten. Allerdings wissen wir schon eine Menge über den Mechanismus, der die Reaktion auslöst, die als Blüte bezeichnet wird. Die Blüte ist nicht das Ergebnis eines einzelnen Phänomens. Es gibt auch nicht das eine „magische“ Hormon, das die Blüte auslöst. Pflanzen blühen in Reaktion auf mehrere Auslöser, die eine ziemlich komplexe Kette von physiologischen und genetischen Reaktionen bilden. Am Ende führt dies zu Veränderungen in den morphologischen Eigenschaften der blühenden apikalen Triebe. Wichtigster Auslöser ist ein bestimmter Effekt des Lichts, den man unter dem Begriff Fotoperiodismus kennt.
Fotoperiodismus steht für die Reaktion einer Pflanze auf bestimmte Lichtsignale. Dabei spielen sowohl die Dauer als auch die Qualität des Lichts, dem die Pflanze ausgesetzt ist, eine wesentliche Rolle. Pflanzen nehmen Licht anders wahr als Menschen oder Tiere. In Pflanzen wirkt der Teil des elektromagnetischen Spektrums, den wir als Licht wahrnehmen, als Energielieferant für bestimmte fotochemische Reaktionen, die bestimmte Prozesse steuern und selbst Energie produzieren. Auch Tiere nutzen Lichtenergie, um ihre Umgebung zu „sehen“. Licht ist eine Dualität: Es existiert gleichermaßen als Elementarteilchen (Foton) und als Welle. Je höher die Frequenz (kürzere Wellenlänge), desto höher der Energiezustand des als Foton bekannten Quantenbündels (siehe Abb. 1). Die fotochemischen Systeme in Pflanzen sind darauf ausgelegt, bestimmte Lichtfrequenzen einzufangen und die Lichtenergie zur Durchführung von chemischen Reaktionen zu nutzen.
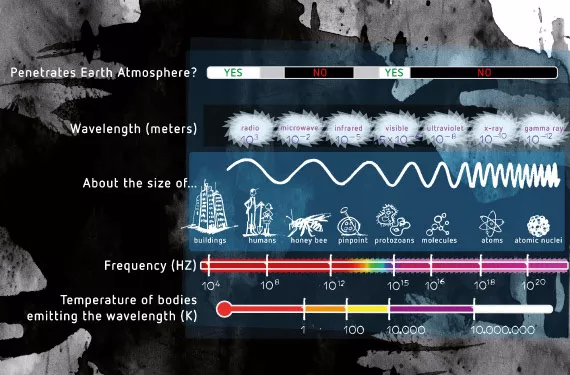
Spektrumfarben
Pflanzen nehmen Lichtenergie grundsätzlich aus zwei Gründen auf: zur Bildung von Kohlehydraten, und zur Regelung lebenswichtiger Prozesse, die sich in den Pflanzenzellen vollziehen. In diesem Artikel nehmen wir Bezug auf die Prozesssteuerung, wobei die Wellenlängen zur Bildung von Kohlehydratenim Grunde ähnlich sind. Folgende 4 Farben des Farbspektrums, die für Pflanzen von Bedeutung sind:
- UV (Ultraviolett) von 340–400 Nm
- Blau von 400–500 Nm
- Rot von 600–700 Nm
- Fernrot (der Anfangsbereich von Infrarot) von 700–800 Nm
Lichtsammelpunkte
Diese Zahlen sind nicht als absolut zu betrachten, weil sich die Farben eigentlich überlappen. Eine Pflanze kann etwas Energie aus dem Bereich 500–600 Nm nutzen, allerdings nicht viel. Die Pflanzen nutzen Pigmente, um die verschiedenen Energie-Wellenlängen einzufangen. Die vier Wellenbereiche der elektromagnetischen Energie steuern die Aktivität der Pflanze über drei Sammelpunkte, auch lichtabsorbierende Pigmente genannt:
- Cryptochrome (blau und UV)
- Phytochrome (rot und fernrot)
- Fototropine (blau und UV)
Die Lichtsammelpunkte fungieren als Schalter, die bestimmte Prozesse in der Pflanze „ein- und ausschalten“ und andere regulieren. Während ein Mensch nur die reflektierten Farben (Wellenlängen oder Frequenz) wahrnimmt und eine Aufhellung oder Dämpfung des Lichts erfährt, reagieren Pflanzen auch auf die Lichtverschiebung zwischen Frequenzen, die sich uns als Intensität zeigt.
Pflanzen, die im Schatten anderer Pflanzen wachsen, erhalten mehr rotes und fernrotes Licht und weniger blaues Licht. Sie reagieren empfindlich auf die Verschiebung von rotem zu blauem Licht, die von Natur aus bei Sonnenaufgang auftritt, sowie auf die entgegengesetzte Verschiebung bei Sonnenuntergang. Auch Änderungen der Zeiten, zu denen diese täglichen Ereignisse stattfinden, werden empfindlich wahrgenommen. Die einzelnen Pigmente fungieren als Schalter, die durch die Energie einer bestimmten Wellenlänge als Verhältnis von einer Frequenz zur anderen betätigt werden. Sogar die Abwesenheit von Licht beeinflusst die Reaktion der Pflanze über diese Steuerzentralen. All diese Steuermechanismen wirken sich auf den Prozess aus, den wir als Blüte kennen.
Licht steuert die natürlichen Rhythmen der Pflanze, so wie es beispielsweise auch den Schlafrhythmus von Tieren steuert. Diese natürlichen Rhythmen, auch zirkadiane Rhythmen, genannt, sind in allen Lebensformen enthalten. Es gibt Zeiten der Aktivität in denen Energie verbraucht wird und Zeiten der Ruhe. All diese Aktivitäten sind in einem Zeitraum einprogrammiert, der mehr oder weniger 24 Stunden umfasst.
Es ergibt wenig Sinn, in der Dunkelheit Chemikalien zu produzieren, die zum Einfangen von Fotonen dienen (obwohl manche Pflanzen dies durchaus tun). Wie bei einer Fabrik müssen Komponenten genau dann eintreffen, wenn sie benötigt werden oder es gibt ein Vorratslager mit einer Mindestmenge. Und die Maschinen müssen laufen, wenn die richtigen Teile bereitstehen. Licht bestimmt diese Rhythmen nicht nur durch seine Präsenz, sondern auch durch seine Qualität.
Eine Pflanze nimmt sowohl die Qualität als auch Quantität des auf sie einstrahlenden Lichts wahr. Je nach Umweltfaktoren wie Luftqualität oder Jahreszeit nimmt die Pflanze unterschiedliche Farbverhältnisse. wahr. Dieser Unterschied wird grundsätzlich von den Pigmenten gemessen, die - im Zusammenwirken mit anderen Auslösern und Prozessen - steuern, wie und wann die Pflanze reagiert. Das Licht stellt die biologische Uhr in der Pflanze, damit alle pflanzeninternen Prozesse harmonisch ablaufen können.
Cryptochrome nehmen die Richtung des Lichts und dessen Quantität wahr. Zu den von Cryptochromen gesteuerten Reaktionen gehören:
- die Funktion der Stomata
- Gentranskription und -aktivierung
- Hemmung der Stängeldehnung
- Pigmentsynthese und
- die Ausrichtung der Blätter auf den Sonnenstand
Fototropine, die anderen Blaulichtrezeptoren, sind als Schadensvermeidungssystem für den Fototropismus oder die Bewegung der Pflanzen und die Bewegung der Chloroplasten innerhalb der Zelle in Reaktion auf die Lichtmenge verantwortlich. Es gibt ebenfalls Anzeichen dafür, dass sie die Schutzzellen an den Stomataöffnungen aktivieren.

Phytochrome: Pr und Pfr
Ein Phytochrom ist ein Komplex von Pigmenten, wobei grundlegend 2 Typen zu unterscheiden sind:
- reagiert auf rotes Licht (Pr)
- reagiert auf fernrotes Licht (Pfr)
Je nachdem, welche Lichtfrequenzen sie am meisten absorbieren (auch wenn die andere Frequenz und das Blaulicht es ebenfalls aktivieren wird). Die beiden Pigmente wandeln sich im Allgemeinen ständig vom einen Typ zum anderen: Pr wird mit Rotlicht zu Pfr und umgekehrt (obwohl manche Pr/Pfr-Formen je nach Lichtertrag, Intensität und Lichtqualität ihre Fähigkeit zur Umwandlung verlieren). Die aktive Form, die Reaktionen wie die Blüte auslöst, ist Pfr. Rotes Licht übt den größten Einfluss auf die Fotomorphogenese (die Wirkung von Licht auf die Pflanzenentwicklung) aus. Fernrotes Licht kann wiederum manche Pfr-Reaktionen umkehren.
Das Phytochrom steuert viele Funktionen wie z. B.
- Genexpression und -repression
- Gentranskription
- Dehnung von Sämlingen und Stängeln
- Keimung
- Fotoperiodismus (die Blütenreaktion)
- Schattenvermeidung und Anpassung an unterschiedliche Lichtstärken
- Chlorophyllsynthese

Ein Beispiel für eine Rotlichtreaktion ist die Änderung des Lichtintervalls von langen auf kurze Tage, was in Kurztagpflanzen die Blüte auslöst.Das liegt daran, dass die Pflanze die Veränderung durch den Verhältnisunterschied zwischen rotem und fernrotem Licht (oder überhaupt keinem Licht) spürt und anfängt, ihre Physiologie von einem Zustand des vegetativen Wachstums in einen Zustand des floralen Wachstums zu verändern. Wenn die Pflanze Licht erhält, ist das Pr/Pfr-Verhältnis (Pr: Pfr) ungefähr im Gleichgewicht (de facto ist Pfr leicht höher).
Pr wird während der Dunkelheit auf natürliche Weise von der Pflanze produziert und häuft sich an. Pfr baut sich ebenfalls langsam zu Pr ab (die Halbwertszeit beträgt ca. 2,5 Stunden). Am nächsten Morgen, bei völliger Helligkeit, ist das Pr/Pfr-Verhältnis wieder ausgeglichen. Man könnte Pfr mit den Sandkörnern in einer Sanduhr vergleichen. Derzeit geht man davon aus, dass Kurztagpflanzen bei niedrigen Pfr- und hohen Pr-Konzentrationen blühen, Langtagpflanzen jedoch nicht. Wenn die Konzentrationen höher und die Pr-Konzentrationen geringer sind, blühen Langtagpflanzen im Gegensatz zu Kurztagpflanzen.
Langtag und Kurztag
Wenn wir zwei Pflanzen betrachten, bei denen die eine auf eine Blüte bei einer Tageslänge von 10 Stunden bei 14-stündiger Dunkelheit ausgelegt ist (Kurztagpflanze) und die andere bei einer Tageslänge von 14 Stunden und 10-stündiger Dunkelheit blüht (Langtagpflanze), dann ist die Nacht ausschlaggebend für die Blüte. Dieser Prozess wird in Abb. 3 veranschaulicht. Tatsächlich benötigt die Kurztagpflanze 14 Stunden Dunkelheit, um Pr zu akkumulieren. Auch muss genügend Pfr in Pr umgewandelt werden, damit das Pfr-Niveau über Nacht so lange unterdrückt wird, dass eine morphologische Veränderung möglich wird. Diese Änderung wird nach einer bestimmten Anzahl von Tagen nicht umkehrbar (irreversibel). In einer Langtagpflanze ist der Prozess analog. Solche Pflanzen reagieren auf stärkere Pfr-Niveaus.

Florigen, das Blütesignal
Florigen, einst als theoretisches Hormon bezeichnet, gilt inzwischen allgemein als Botschafter-RNA und ist unter der Bezeichnung FT mRNA bekannt. Sehr vereinfacht ausgedrückt, handelt es sich hier um ein Proteinmolekül, das auf einem Abschnitt der DNA einer Pflanze in einem Bereich produziert wird, den man als FLOWRING LOCUS T (FT) bezeichnet. Dieses Protein ist wie ein Schlüssel, der nach einem bestimmten Schloss sucht, in welches er hineinpasst. Dreht sich der Schlüssel im Schloss, so werden wiederum andere Prozesse initiiert. In Kombination mit einem anderen Gen, das als CONSTANS (CO), bezeichnet wird, gilt inzwischen als erwiesen, dass dies die Veränderung vom vegetativen Status zum Blütestatus auslöst. Die Veränderung einer Pflanze zur Blüte benötigt demnach externe Signale, die die Prozesse der Pflanze beeinflussen, steuern und die die Genexpression auslösen. All dies wird durch die Lichtveränderungen, die von der Pflanze wahrgenommen werden, ausgelöst.
Blütenreaktion
Grundsätzlich gibt es 5 Arten von Blütenreaktionen in Pflanzen.
- Kurztagpflanzen (KTP), die nur eine Verlagerung hin zu kurzen Tagen und langen Nächten benötigen, um blühen zu können
- Langtagpflanzen (LTP), benötigen wiederum genau das Umgekehrte
- Langkurztagpflanzen (LKTP)...
- ... und Kurzlangtagpflanzen (KLTP), die eine bestimmte Zeit als Lang- bzw. Kurztagpflanze, gefolgt von einem kurzen bzw. langen Tag, benötigen, um blühen zu können.
- Die tagesneutralen Pflanzen (TNP), die zwar dieselben Lichtfunktionen benötigen, bei denen jedoch andere Faktoren (d. h. nicht die Tageslänge) die Blüte auslösen.
In allen Fällen ist es nicht nur die Art oder Qualität des einstrahlenden Lichts, die die Blüte auslöst, sondern auch die Dauer des Lichteinfalls (Ausnahme: TNP).
Genau genommen ist es die Dauer der Abwesenheit des Lichts zur Nachtzeit, die die Blüte auslöst, allerdings gestützt auf Prozesse und Metaboliten (Pfr etc.), die vom Licht gelenkt werden. Man sollte sich vor Augen führen, dass es neben den hier beschriebenen Prozessen vermutlich viele weitere Prozesse gibt, die hier eine Rolle spielen, z. B. die Wechselwirkung anderer Gene und Hormone wie GA (Gibberellinsäure)..
Licht ist entscheidend für alles Leben, vor allem aber für die Pflanzenwelt. Dort produziert das Licht nicht nur die Substrate für Wachstum und Stoffwechsel, sondern gibt auch die Rhythmen und Zyklen der täglichen Routine vor. Licht steuert entscheidende Aspekte des Überlebens und der Vermehrung: Es gibt den Takt für das Leben in sämtlichen Organismen vor. Eine weitere, genauso wichtige Erkenntnis: Licht ist nicht immer gleich Licht, zumindest was Pflanzen betrifft. Die korrekten Lichtverhältnisse (Blau zu Rot, Rot zu Fernrot usw.) müssen zur Verfügung stehen, damit eine Pflanze existieren kann. Wie bei allen anderen Dingen kann es auch für eine Pflanze manchmal zu viel des Guten sein. Und obwohl Licht absolut lebenswichtig für Pflanzen ist - am Ende ist es doch nur einer von vielen Faktoren, die das Leben einer Pflanze und das Leben an sich prägen.
Quellenangaben
- Aushulz. "file:Chlorophyl_ab_spectra2.png." Wikipedia. http://en.wikipedia.org/wiki (accessed 4 10, 2011).
- Huang, T., Bōhlenius, H., Eriksson, S., Parcy, F., Nilsson, O. "The mRNA of the gene FT moves from leaf to shoot apex and induces flowering." Science 309 (2005): 1694-1696.
- Kasperbauer, M. J., and D. L. Karlen. "Plant Spacing and Reflected Far-Red Light Effects on Phytochrome-Regulated Photosynthate Allocation in Corn Seedlings." Crop Sci. 34 (1994): 1564-1569.
- Koning, Ross E. Photoperiodism. 1994. http://plantphys.info/plant_physiology/photoperiodism.shtml (accessed May 31, 2011).
- Taiz, L., and E. Zeiger. Plant Physiology. 3rd. Sunderland: Sinauer Associates, Inc., 2002.
- Zeevart, Jan A.D. "The Plant Cell 18:1783-1789 (2006)." The Plant Cell. August 2006. http://www.plantcell.org/content/18/8/1783.full#BIB18 (accessed June 9, 2011).

